The southeastern United States harbors the richest freshwater fish fauna on the North American continent north of Mexico (Burr and Mayden, 1992), but portents of decline of this great fauna are increasingly acknowledged (e.g., Burr and Warren, 1986; Mount, 1986; Burkhead and Jenkins, 1991; Etnier and Starnes, 1991; Ross and Brenneman, 1991; Gilbert, 1992; Warren and Burr, 1994; Angermeier, 1995; Menhinick, in press). Southeastern fishes (493 species) comprise about 47 percent of the North American fish fauna (1,061 species) and 62 percent of the fauna in the United States (790 species) (Page and Burr, 1991; Burr and Mayden, 1992). Within the United States, imperilment of southeastern fishes is second only to that of western fishes (Williams et al., 1989; Minckley and Deacon, 1991; Warren and Burr, 1994). Unlike the southeastern fish fauna, the western fish fauna of North America is relatively depauperate, and the proportion of the western fish fauna that is extinct or threatened with extinction is high (Miller et al., 1989; Moyle and Williams, 1990; Warren and Burr, 1994). In response to wholesale losses of native fishes, comprehensive programs in the west are underway or planned to conserve and recover fish diversity (e.g., Minckley and Deacon, 1991; Moyle and Yoshiyama, 1994), but success of these reactive approaches are as yet unknown. Increasing recognition of the decline of fishes and aquatic habitats in the Southeast, both harbingers of the western situation (Minckley and Deacon, 1991), should be the clarion call for proactive efforts toward conservation of the richest fish fauna in the United States.
Known extinctions of southeastern fishes are limited to two species, the harelip sucker (Moxostoma lacerum) and whiteline topminnow (Fundulus albolineatus) (Miller et al., 1989), but reduction in range, extreme isolation of extant populations, and extirpation of fishes from entire drainages are common (e.g., Etnier et al., 1979; Burr and Page, 1986; Starnes and Etnier, 1986; Jenkins and Burkhead, 1994; Warren et al., 1994). Fish extinction or local extirpation is nearly always attributable to multiple human impacts, and cumulative effects from complex aquatic degradation may obscure association of geographic, population, or ecological characteristics with extinction or extirpation patterns (Miller et al., 1989; Moyle and Leidy, 1992; Frissel, 1993; Angermeier, 1995).
Successful management for maintenance of fish diversity in the Southeast is, as in the West, a battle against extinction (Minckley and Deacon, 1991) and ultimately, a battle for ecological integrity at landscape scales (Angermeier and Karr, 1994). Studies of fishes (Sheldon, 1987; Etnier and Starnes, 1991; Nagel, 1991; Angermeier, 1995), island birds (Terborgh and Winter, 1980; Karr, 1990), and meta-population dynamics (Hanski, 1982) relate the process of extinction to factors that decrease habitat area and increase insularization (Angermeier, 1995). However, loss of diversity via extinction is not usually observable nor cataclysmic. Rather, the process is incremental with total extinction preceded by local or regional extirpations that usually reflect a populations sensitivity to decreasing habitat area and increasing isolation (Angermeier, 1995). Recent state and regional analyses of extirpation patterns (Sheldon, 1987; Etnier and Starnes, 1991; Nagel, 1991; Moyle and Leidy, 1992; Frissel, 1993) indicate that landscape-scale phenomena such as decreasing habitat area and increasing habitat fragmentation are associated strongly with regional loss of fish diversity. Theoretical considerations suggest that local extinction is accelerated when landscapes are insularized if meta-population dynamics are important (Hanski, 1982). Meta-population concepts have been applied only recently to stream fish conservation issues (Schlosser and Angermeier, 1995). Understanding (and ultimately preventing) anthropogenic extinction is likely to require greater focus on landscape-level patterns and processes than in traditional conservation approaches.
The Clean Water Act of 1972, as amended, is a potentially powerful statutory vanguard for proactive management for ecological integrity of aquatic systems (Blockstein, 1992; Angermeier and Karr, 1994), but the last line of defense against extinction of fishes in the United States is the Endangered Species Act of 1973, as amended. To effectively implement existing laws, two primary types of information are needed in the conservation arsenal (Warren and Angermeier, in press): accounting and ecological information. Accounting information (e.g., the presence or absence of taxa or communities) comes from inventories of sites, watersheds, or ecoregions. These data are necessary to integrate levels and geographic distributions of fish diversity across multiple spatial scales and to identify fishes and fish communities that are unique, rare, or imperiled. Ecological information, such as habitat needs or evolutionary history, helps elucidate factors involved in the generation and maintenance of fish diversity, may help predict potential losses of fish diversity, enhances successful recovery of diminished fish diversity, and identifies taxa or communities that are sensitive indicators of ecological integrity. The availability and synthesis of these two types of information are requisite to elucidating landscape-level patterns and developing effective management for native fishes.
Basic accounting information is available for southeastern fishes but has not been generally synthesized or analyzed for conservation assessment or planning (Etnier and Starnes, 1991; Angermeier, 1995; Warren and Angermeier, in press). Although ecological information also is available, the quality of that information varies widely for different fishes. As such, we rely here primarily on accounting information to discern patterns of imperilment at landscape scales. To date, there has been little effort to examine large-scale patterns of diversity and imperilment of southeastern fishes with the objectives of discovering general principles underlying imperilment that may be useful in proactive management or conservation triage (Frissel, 1993; Angermeier, 1995).
We provide here a beginning toward the large-scale synthesis of accounting and to a limited extent ecological information for fishes of the southeastern United States. In doing so, we present an up-to-date, comprehensive inventory of fishes of the Southeast and use geographical displays of fish and stream diversity and imperilment to convey the richness, spatial extent, and variation in these characteristics. For individual fishes and fish families, we ask two questions: is range size associated with imperilment, and is imperilment a function of familial membership? For major river drainages of the Southeast, we pose three questions: is fish imperilment associated with drainage area, native fish taxa richness, endemism, or stream-type diversity; which of these variables are the best predictors of imperilment; and what are the implications of the identified predictors?
Our specific objectives are to provide an updated distributional checklist of all southeastern freshwater fishes; summarize geographical patterns of fish imperilment, fish diversity, and stream diversity by state and major rivers in the southeastern United States; and examine relationships of numbers of imperiled taxa to native fish taxa richness, geographic range, drainage area, and stream-type diversity. We believe the maps and accompanying analyses are useful initial steps in prioritizing and coordinating conservation actions for fishes and other aquatic resources in the Southeast and in highlighting the urgent need for holistic approaches to aquatic conservation.
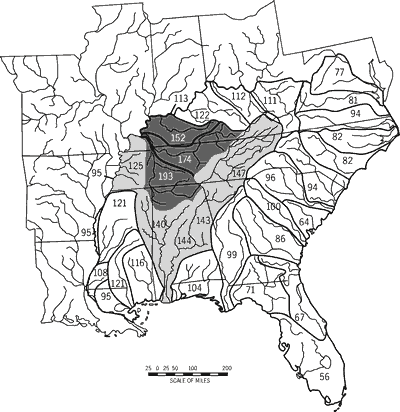
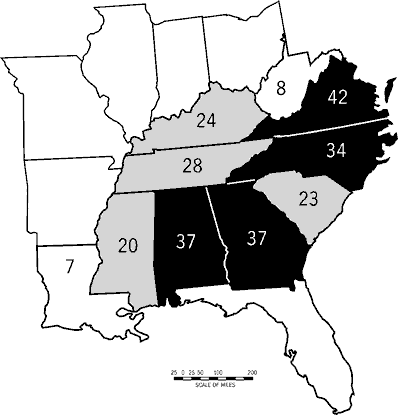
Our study area, referenced as the Southeast or southeastern United States, includes Alabama, Florida, Georgia, Kentucky, Louisiana, Mississippi, North Carolina, South Carolina, Tennessee, Virginia, and West Virginia (Figure 1). We followed Warren and Burr (1994) for the number of native fishes in each state. Although these totals may not exactly match those of others, they do illustrate patterns in fish diversity and levels of imperilment among southeastern states (Warren and Burr, 1994).
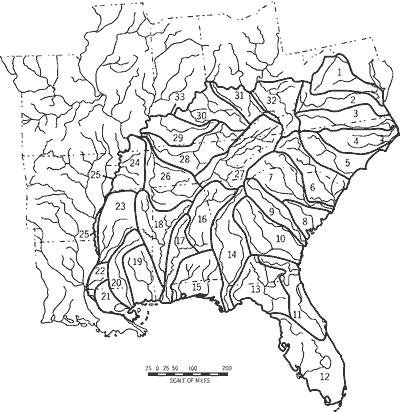
Within the 11-state study area, we recognized 33 drainage units (DU1-DU33) grouped into 11 regions (A-K) (Table 1; Figure 2). We delimited drainage units and regions based upon fish faunal similarity analyses (Burr and Warren, 1986; Hocutt et al., 1986; Swift et al., 1986; Warren et al., 1991), vicariance biogeography analyses (Mayden, 1988), drainage propinquity and interconnectivity, and debauchment into a common lake, sound, or bay (e.g., Chesapeake Bay, DU1-DU2; Albemarle and Pamlico sounds, DU3-DU4; Lake Pontchartrain, DU21).
As a measure of stream-type diversity, we calculated the number of hydrologic-physiographic types for each drainage unit and state in the study area. Each "stream type" is a unique combination of three attributes: drainage unit, stream size, and physiography. Drainage units are defined in Table 1. Using a 1:106-scale map (Fenneman and Johnson, 1964), we recognized three stream sizes: <_fourth order, fifth through seventh order, and >_ eighth order. We used six physiographies (Fenneman and Johnson, 1964): Coastal Plain, Piedmont, Blue Ridge, Ridge and Valley, Appalachian Plateau, and Interior Low Plateaus. We took drainage-unit areas (n = 30) from stream gauging station compendia (Anderson, 1950; Hains, 1968) and Burr and Warren (1986), Swift et al. (1986), and Jenkins and Burkhead (1994). We did not estimate areas for DU12, DU25, or DU33.
1) Potomac-Rappahannock-York River Drainages
2) James River Drainage
3) Roanoke River Drainage (including Chowan River)
4) Tar-Neuse River Drainages
5) Cape Fear River Drainage (including coastal drainages from Cape Lookout to mouth of Cape Fear River)
6) Peedee River Drainage (from mouth of Cape Fear River to and including Peedee River)
7) Santee-Cooper River Drainages (from mouth of Peedee River to mouth of Cooper River)
8) Edisto-Combahee River Drainages (from mouth of Cooper River including Combahee and Coosahatchie systems to mouth of Savannah River)
9) Savannah River Drainage
10) Ogeechee-Altamaha River Drainages (from mouth of Savannah River to and including Altamaha River)
11) Satilla-St. Marys-St. Johns River Drainages (from mouth of Altamaha River to and including St. Johns River)
12) Everglades-Tampa Bay-Waccasassa River Drainages (northwest to and including Waccasassa River)
13) Suwannee-Aucilla-Ochlockonee River Drainages (from mouth of Waccasassa River to Apalachicola Bay)
14) Apalachicola Basin (including Chipola, Chattahoochee, and Apalachicola rivers)
15) St. Andrew-Choctawhatchee-Pensacola Bay Drainages
16) Coosa-Tallapoosa River Systems
17) Lower Alabama-Cahaba River Systems (including Mobile Bay)
18) Tombigbee-Black Warrior River Systems
19) Pascagoula-Biloxi-Bay St. Louis Drainages (from Mobile Bay including Escatawpa and Bay St. Louis systems to mouth of Pearl River)
20) Pearl River Drainage
21) Lake Pontchartrain Drainage (from mouth of Pearl River)
22) Minor Mississippi Tributaries South (eastern tributaries from mouth of North River to mouth of Black River)
23) Black-Yazoo River Systems
24) Minor Mississippi Tributaries North (from mouth of Yazoo River to and including Mayfield Creek)
25) Mississippi River Mainstem
26) Lower Tennessee River System (from mouth Sequatchie River downstream)
27) Upper Tennessee River System (from and including Sequatchie River upstream)
28) Cumberland River System
29) Green-Tradewater River Systems (from Mayfield Creek to mouth of Green River)
30) Kentucky-Salt River Systems (from mouth of Green River to mouth of Licking River)
31) Licking-Big Sandy River Systems (from and including mouth of Licking River to mouth Guyandotte River)
32) Kanawha-New-Guyandotte River Systems (from and including Guyandotte to Kanawha-New)
33) Ohio River Mainstem
We obtained presence or absence and native versus non-indigenous status of fishes within a particular drainage unit (Appendix 1 [PDF--63 KB]) from Lee et al. (1980), Hocutt et al. (1986), Starnes and Etnier (1986), and Swift et al. (1986). We updated this information from distribution maps in Burr and Warren (1986), Page and Burr (1991), Ross and Brenneman (1991), Etnier and Starnes (1993), and Jenkins and Burkhead (1994). We obtained distribution information concerning species described subsequent to the previously cited works, those resurrected from synonymy, and subspecies elevated to species from the following sources: Bauer et al. (1995, Etheostoma scotti); Boschung et al. (1992, Etheostoma chermocki); Burr and Page (1993, Percina stictogaster); Gilbert et al. (1992, Fundulus auroguttatus and F. rubrifrons); Mayden (1993, Elassoma alabamae); Page et al. (1992, Etheostoma chienense, E. pseudovulatum, E. oophylax, E. corona, and E. forbesi); Suttkus (1991, Notropis rafinesquei); Suttkus and Etnier (1991, Etheostoma tallapoosae and E. brevirostrum); Suttkus and Bailey (1993, Etheostoma colorosum and E. bellator); Suttkus et al. (1994a, Etheostoma raneyi, E. lachneri, and E. ramseyi); Suttkus et al. (1994b, Percina aurora and P. brevicauda); Thompson (1995, Percina austroperca); Warren (1992, Lepomis miniatus); Warren et al. (1994, Notropis albizonatus); Williams and Clemmer (1991, Scaphirhynchus suttkusi); and Wood and Mayden (1993, Etheostoma douglasi, E. chuckwachatte, and E. etowahae). For native fishes, we followed the familial arrangement, common names, and nomenclature of Mayden et al. (1992), and for exotic species, Page and Burr (1991). In the checklist (Appendix 1 [PDF--63 KB]), we arranged genera and species alphabetically within families.
"Unique taxa" and "range extent" are indices of endemicity and cosmopolitanism, respectively, across the Southeast, as judged from known historical distributions of extant native fishes. Unique taxa are those restricted to one drainage unit. Most of these are endemic to one drainage unit in the Southeast, but a few are peripheral and occur outside the area. Range extent is the number of drainage units in which a particular taxon occurs or historically occurred.
We took the conservation status of fishes from lists published by the American Fisheries Society (AFS) (Deacon et al., 1979; Williams et al., 1989) and U.S. Fish and Wildlife Service (1994a, 1994b) (Appendix 1 [PDF--63 KB]). Reference to "imperilment" or "imperiled" includes those taxa with any one of three AFS conservation status categories as recognized by Williams et al. (1989). We used this definition of imperilment for statistical analyses because of the time lag between recognition of a species as being imperiled and actual protection under the Endangered Species Act of 1973, as amended (Reffalt, 1991; Warren and Burr, 1994). The AFS categories included endangered (AFS-E), threatened (AFS-T), and of special concern (AFS-SC). We also used two combinations of these, AFS-ET and AFS-ETSC, in analyses to distinguish among: 1) the most critically imperiled fishes and 2) all fishes with a conservation status, respectively. Reference to "federal status categories" included endangered (E), threatened (T), proposed endangered (PE), proposed threatened (PT), candidate 1 (C1), and candidate 2 (C2) as listed in U.S. Fish and Wildlife Service (1994a, 1994b). "Protected federal categories" included only the first four of these (Littell, 1992).
We calculated "percentage imperilment" as the number of imperiled fishes in a drainage unit divided by the total number of native fish taxa in that unit × 100. We estimated percentage increase in imperilment as the total number of imperiled fish taxa in a drainage unit minus the total number of taxa recognized in Deacon et al. (1979) divided by the native fish taxa richness in the unit × 100. As ostensible temporal rates, we recognize these estimates do not account for the discovery of new or better distribution information for fishes nor the discovery of previously unrecognized taxa. As such, the assumption of a linear increase in imperilment over the ten-year interval may be invalid. The Deacon et al. (1979) and Williams et al. (1989) papers remained, however, the only temporal benchmarks available from which estimates could be made.
We calculated familial imperilment, expressed in percent, as the number of imperiled taxa (AFS-ETSC) in a family divided by the number of native taxa represented in that family in the Southeast. We tested independence of total imperiled taxa in a family and familial taxa richness for the six most taxa-rich families using the likelihood ratio test (LI) (StatXact-Turbo, Mehta and Patel, 1992; G-test equivalent, Sokal and Rohlf, 1981). To help insure that no variables with potentially useful associative or predictive value were overlooked, we conducted all hypothesis testing at p < 0.10 (see Angermeier, 1995 and papers cited therein).
We assessed the relationship between range extent and imperilment across fish taxa using the Kruskal-Wallis test, analysis of variance with a priori orthogonal contrasts of means, and logistic regression (SAS Institute, Inc., 1994). For the Kruskal-Wallis test and analysis of variance, class variables were AFS-E, AFS-T, AFS-SC, and no status; the response variable was range extent. For logistic regression, imperilment (AFS-ETSC) and no status were the response variables, and range extent, the independent variable. We evaluated goodness of fit of the logistic model following Hosmer and Lemeshow (1989).
We tested total native taxa richness, imperilment (AFS-ET and AFS-ETSC), unique taxa, and stream-type diversity across the 33 drainage units for the departure of spatial distribution from random using the Index of Dispersion (ID) where: ID = s2(n - 1)/x_ and s2 is the variance; n, the sample size; and x_ is the mean (Southwood, 1978). The index is distributed approximately as c2 and approaches zero for samples from a random spatial distribution (i.e., Poisson distribution). Conversely, a large ID implies spatial aggregation or clumping.
Across drainage units, we examined relationships of imperilment (AFS-ET and AFS-ETSC) to drainage-unit area, unique taxa, stream-type diversity, and total native taxa richness using correlation and multiple regression analyses in SAS Institute, Inc. (1994). We used both nonparametric and parametric correlation coefficients (Kendalls tau-b and Pearsons, respectively). We used Type II sums of squares to assess relative predictive capabilities of several independent variables for imperilment (Angermeier and Schlosser, 1989). After examination of scatterplots, we log-transformed drainage-unit area, imperilment, and native taxa richness as log10(x) or log10(x + 1) to minimize effects of non-linearity (Sokal and Rohlf, 1981).
We compiled a comprehensive list of 530 freshwater fish taxa for the southeastern United States (Appendix 1 [PDF--63 KB]). This inventory included 496 extant native taxa (493 species) and 34 non-indigenous, established fishes. Interdrainage transfers of indigenous fishes by humans also are common in the Southeast. At least 62 indigenous southeastern fish species, 13 percent of the fauna, have been introduced to drainages in the Southeast to which they are not native.
Eighty-four fish taxa in the Southeast are recognized as imperiled (Table 2). Fifty taxa (ten percent of the fauna) are placed in AFS-E and AFS-T categories and 34 (seven percent) are recognized as AFS-SC. Thirty-two fishes (six percent of the fauna) are in protected federal categories and an additional 52 (11 percent) are candidate species. Although total numbers recognized in federal status categories versus AFS status categories are equal, the AFS assessment differs notably in the number of fish species considered threatened. By either source about 17 percent of the southeastern fish fauna is considered in need of conservation actions.
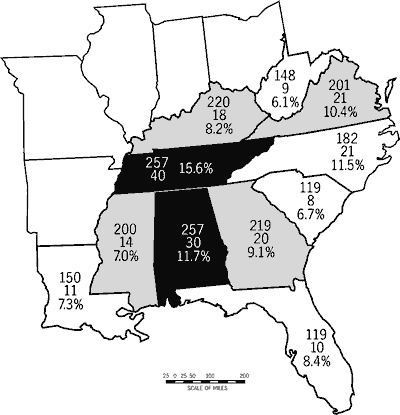
The southeastern states have a high diversity of fishes, moderate to high imperilment, and a wide range of stream-type diversity. Six southeastern states have native freshwater fish faunas of 200 or more taxa (Figure 1). Tennessee and Alabama are centers of diversity; each harbors at least 257 native fish taxa. Kentucky and Georgia follow with 220 and 219 fishes, respectively. No southeastern state has fewer than 119 fish taxa.
Nine of the 11 southeastern states support ten or more imperiled fishes (Figure 1). Alabama and Tennessee have the highest number of imperiled fishes (30 and 40 taxa, respectively) followed by Georgia, Kentucky, North Carolina, and Virginia (18-21 fishes). Percentage imperilment is highest in Tennessee (15.6 percent), is greater than ten percent in Alabama, North Carolina, and Virginia, and is lowest in West Virginia (6.1 percent).
The Southeast lies within several major physiographic regions and is drained by numerous large rivers. These factors are reflected in the tally by state for stream-type diversity. Stream-type diversity ranges from seven stream types in Louisiana to 42 in Virginia. Georgia, Alabama, and North Carolina also show high stream-type diversity (>_34; Figure 3).
Table 2. Federal and American Fisheries Society (AFS) conservation status category totals for fishes of the southeastern United States. |
||
Status Categories |
Federal |
AFS |
Endangered |
18 |
14 |
Proposed Endangered |
1 |
- |
Threatened |
13 |
36 |
Proposed Threatened |
- |
- |
Subtotal |
32 |
50 |
Candidate |
52 |
- |
Special Concern |
- |
34 |
Total |
84 |
84 |
Imperilment is not distributed evenly among fish families in the Southeast. Of 30 families of native fishes, 12 have one or more imperiled members (Table 3). Among these 12 families, percent imperilment is variable, ranging from seven percent regarding the sunfishes (Centrarchidae) to 86 percent regarding the sturgeons (Acipenseridae). High percent imperilment also occurs in the pygmy sunfishes (Elassomatidae) and cavefishes (Amblyopsidae), with 50 percent and 40 percent (respectively) of family members imperiled.
Among the six most taxa-rich families, imperilment status (AFS-ETSC versus no status) and family membership are associated (LI = 30.41, p < 0.0001, 5 df) (Table 4). The perches and darters (Percidae, primarily the genera Ammocrypta, Etheostoma, and Percina) and bullhead catfishes (Ictaluridae, mostly madtoms of the genus Noturus) have more imperiled members than expected based on their representation in the fauna; the suckers (Catostomidae) are imperiled in approximate proportion to faunal representation; and the minnows (Cyprinidae), topminnows (Fundulidae), and sunfishes (Centrarchidae) have fewer imperiled taxa than expected (Table 4).
Imperilment is related negatively to range extent for native fishes in the Southeast. Both the Kruskal-Wallis test and analysis of variance showed significant differences in range extent among status categories (c2 = 9.08, p < 0.0282; F = 3.674, p < 0.0122, respectively). Orthogonal contrasts of the means indicated range extent of imperiled taxa is significantly lower than non-imperiled taxa (F = 10.63, p < 0.0012)(Table 5). Contrasts did not reveal significant differences in range extent among imperiled taxa (AFS-SC versus AFS-ET, F = 1.17, p < 0.28; AFS-E versus AFS-T, F = 0.47, p < 0.47), but mean range extent was distributed along an increasing gradient from AFS-E to AFS-SC (Table 5).
Probability of imperilment increased with decreasing range extent. Logistic regression, modeling the probability of imperilment on range extent, yielded a significant model with a good fit (a = -1.16, Walds c2 = 9.07, p < 0.0026; b = -0.06, Walds c2 = 50.15, p < 0.0001; Hosmer and Lemeshow c2 = 4.97, p = 0.3855). Calculations from estimates of model parameters (following Rita and Ranta, 1993) indicated probability of imperilment increases from > 0.1 to > 0.2 as range extent decreases from < 16.2 to < 3.5 drainage units.
Fish Taxa Richness
Fish diversity among the 33 drainage units was less variable than any other attribute examined and was not distributed randomly (Table 6). Drainage units with the highest fish species richness (>150 taxa) formed a geographically contiguous core of diversity (Figure 4; Appendix 1 [PDF--63 KB]) that included the Lower Tennessee River System (DU26), Cumberland River System (DU28), and Green-Tradewater River Systems (DU29). A rough semicircle of secondary richness (125 to 147 fishes) is formed to the south (Mobile Basin Region, DU16-DU18), east (Upper Tennessee River System, DU27), and west (Minor Mississippi Tributaries North, DU24) of the richest units.
Table 3. Familial imperilment (i.e., number of imperiled taxa in family; percent of family taxa that are imperiled reported in parentheses), familial taxa richness, and percent familial representation for native fishes of the southeastern United States. Imperiled taxa are those recognized by the American Fisheries Society (see Williams et al., 1989). |
|||
Family |
Familial Imperilment |
Familial Taxa Richness |
Familial Percent of Native Fishes |
Acipenseridae |
6 (86) |
7 |
1 |
Amblyopsidae |
2 (40) |
5 |
1 |
Amiidae |
0 |
1 |
<1 |
Anguillidae |
0 |
1 |
<1 |
Aphredoderidae |
0 |
1 |
<1 |
Atherinidae |
0 |
3 |
<1 |
Catostomidae |
6 (18) |
34 |
7 |
Centrarchidae |
2 (7) |
28 |
6 |
Clupeidae |
0 |
8 |
2 |
Cottidae |
1 (14) |
7 |
1 |
Cyprinidae |
16 (11) |
151 |
31 |
Cyprinodontidae |
0 |
2 |
<1 |
Elassomatidae |
3 (50) |
6 |
1 |
Esocidae |
0 |
3 |
<1 |
Fundulidae |
2 (9) |
22 |
4 |
Gasterosteidae |
0 |
1 |
<1 |
Hiodontidae |
0 |
2 |
<1 |
Ictaluridae |
7 (21) |
33 |
7 |
Lepisosteidae |
0 |
5 |
1 |
Lotidae |
0 |
1 |
<1 |
Moronidae |
0 |
4 |
<1 |
Percidae |
36(24) |
152 |
31 |
Percopsidae |
0 |
1 |
<1 |
Petromyzontidae |
0 |
8 |
2 |
Poeciliidae |
0 |
4 |
<1 |
Polyodontidae |
1(100) |
1 |
<1 |
Rivulidae |
1(100) |
1 |
<1 |
Salmonidae |
0 |
1 |
<1 |
Sciaenidae |
0 |
1 |
<1 |
Umbridae |
0 |
2 |
<1 |
Unique Taxa
Among all 33 drainage units, 128 fishes (26 percent of the native fauna) are unique to a given, single unit. Numbers of unique taxa across drainage units were highly variable spatially and were not distributed randomly (Table 6). A primary center of unique taxa (> 10 unique taxa) was located in the Coosa-Tallapoosa River Systems (DU16) and the Tennessee-Cumberland Region (DU26-28) (Figure 5). A secondary tier of unique taxa (five to seven unique taxa) occurred peripheral to the primary center and included the Tombigbee-Black Warrior River Systems (DU18) and Apalachicola-Florida Panhandle Region (DU14-DU15) to the southwest and east, respectively; the Green-Tradewater River Systems (DU29) to the northwest; and the Roanoke River Drainage (DU3) and Kanawha-New-Guyandotte River Systems (DU32) to the northeast.
Levels of Imperilment
Imperilment was highly variable among drainage units and neither AFS-ET nor AFS-ETSC status combinations were distributed randomly (Table 6). Highest imperilment overlayed the core of drainage units with high numbers of unique taxa and, in part, units of highest taxa richness (Figure 6). The drainage units with highest numbers of unique taxa (Tennessee-Cumberland Region, DU26-DU28; Coosa-Tallapoosa River Systems, DU16) also had the highest percentages of imperiled fishes (16 to 19 taxa, 9.2 to 12.6 percent, respectively)(Figure 6). The Lower Alabama-Cahaba River Systems (DU17) and the Kanawha-New-Guyandotte River Systems (DU32) formed a secondary tier of imperilment. Imperilment was lowest along drainages of the Atlantic and easternmost Gulf slopes but ranged as high as seven taxa and seven percent of drainage-unit taxa richness.
Table 4. Imperilment for the six most diverse fish families in the southeastern United States. Expected values reflect a 17 percent overall imperilment of the southeastern fish fauna. A likelihood ratio test indicated association between status (imperiled vs nonimperiled) and family membership (LI = 30.41, p < 0.0001, 5 df). |
||
Family |
Imperilment |
|
Observed1 |
Expected |
|
Percidae (perches and darters) |
36 (24) |
26 |
Ictaluridae (bullhead catfishes) |
7 (21) |
6 |
Catostomidae (suckers) |
6 (18) |
6 |
Cyprinidae (minnows and carps) |
16 (11) |
26 |
Fundulidae (topminnows) |
2 (9) |
4 |
Centrarchidae (sunfishes) |
2 (7) |
5 |
1 Percent reported in parentheses. |
||
Increases in Imperilment
For all drainage units combined, imperiled species increased from 14.4 to 17.4
percent of total native fishes from 1979 to 1989 (Deacon et al., 1979; Williams
et al., 1989). Percentage increases in imperilment among drainage units are
uneven for the ten-year period, but no unit showed decreases (Figure 7). The
largest increases (greater than three percent) occurred in two geographically
contiguous drainage units, the Coosa-Tallapoosa (DU16) and the Upper Tennessee
(DU27) river systems, both with the highest numbers of imperiled fishes (Figures
6 and 7), and a geographically disjunct unit, the Mississippi River Mainstem
(DU25). Eleven other units showed increases from one to two percent: Roanoke
River Drainage (DU3); Edisto-Savannah-Altamaha Region (DU8-DU10); Everglades-Tampa
Bay-Waccasassa River Drainages (DU12); the remainder of the Mobile Bay Basin
Region (DU17, DU18); Lower Tennessee River System (DU26); and units in the Southeastern
Ohio River Region (DU31-DU33). With the exceptions previously noted, most streams
on the Atlantic and Gulf slopes, the Mississippi Embayment, and the Southeastern
Ohio River Region showed low (less than one percent) or no percentage increases
in imperilment.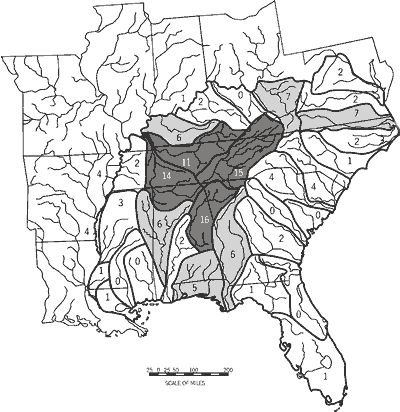
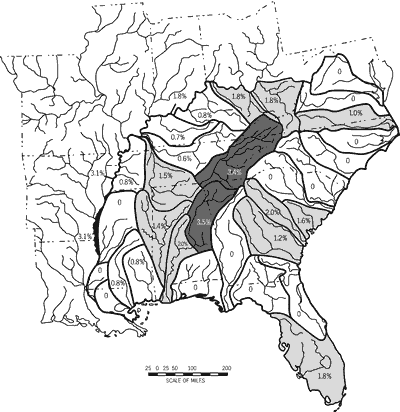
Stream-type Diversity
Stream-type diversity among drainage units was variable over the study area
and was not characterized by abrupt geographic breaks or discontinuities (Figure
8). The departure of the distribution of stream-type diversity from random was
weaker than for native taxa richness, imperilment, or unique taxa (Table 6).
High stream-type diversity occurred in drainage units that arise in uplands
and continue through and/or across lowlands; low diversity occurred in units
primarily restricted to lowlands (Figure 8). Drainage units with the highest
stream-type diversity (nine to ten) included: James River Drainage (DU2); Roanoke
River Drainage (DU3); Coosa-Tallapoosa River Systems (DU16); Potomac-Rappahannock-York
River Drainages (DU1); and the Lower Tennessee River System (DU26). Low stream-type
diversity (>_three) occurred in a geographically and faunistically eclectic
group of drainage units, but with few exceptions these units have relatively
small drainage areas and/or are predominated by lowlands. These units included
the Green-Tradewater River Systems (DU29) and Ohio River Mainstem (DU33); Yazoo-Black-Mississippi
Minor Tributaries Region (DU22-DU24); Pascagoula-Pearl-Pontchartrain Region
(DU19-DU21); St. Andrew-Choctawhatchee-Pensacola Bay Drainages (DU15); Peninsular
Florida Region (DU11-DU13); and the Edisto-Combahee River Drainages (DU8).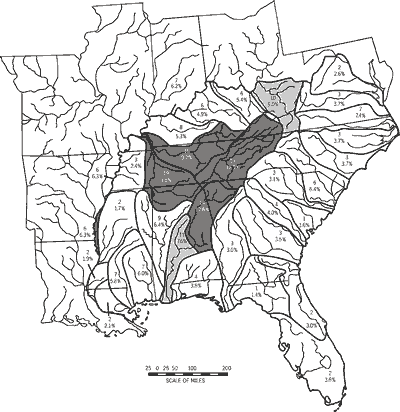
Variables with potential to predict imperilment of fishes in the drainage units included native fish taxa richness, stream-type diversity, drainage-unit area, and unique taxa (Table 7). Native fish taxa richness and unique taxa were correlated significantly with imperilment (AFS-ET and AFS-ETSC). Drainage-unit area and stream-type diversity showed weaker correlations with imperilment.
Because of the tendency for "predictor" variables to be intercorrelated (e.g., stream-type diversity and drainage-unit area; native taxa richness and unique taxa), we used two multiple regression models (Angermeier and Schlosser, 1989) to assess the relative usefulness of native fish taxa richness, stream-type diversity, drainage-unit area, and unique taxa in predicting imperilment (AFS-ET and AFS-ETSC) (Table 8). The first models regressed imperilment on all four variables. Drainage-unit area and unique taxa did not contribute a significant proportion of the sums of squares (Type II SS) for imperilment, and their removal from the models reduced R2 values by < 1.5 percent for both AFS-ET and AFS-ETSC status combinations. The second model included only native taxa richness and stream-type diversity as independent variables. Both variables contributed a significant proportion of the sums of squares for imperilment and were useful predictors for fish imperilment in drainages of the Southeast. In addition, we regressed imperilment on drainage-unit area only, and although the models were significant, R2 values were < 0.15 for both status combinations.
Alabama, Georgia, North Carolina, Tennessee, and Virginia face some of the greatest and most immediate challenges of all southeastern states in future management of native fishes. Each of these states ranks within the top three for two or more of the attributes of fish taxa richness, imperilment, and stream-type diversity (Figures 1 and 3). Nevertheless, we emphasize that all southeastern states support relatively rich fish faunas. The average taxa richness of 188 fishes among southeastern states equals or surpasses that of all other of the lower 48 states except Arkansas and Missouri (Warren and Burr, 1994). Likewise, even states that do not show high values for fish taxa richness, imperilment, or stream-type diversity face critical problems in conservation of native fishes. In Florida (and other Atlantic states), for example, spawning runs are severely curtailed for several anadromous species as a result of dams on large coastal tributaries (Gilbert, 1992). In Mississippi, flood control projects and short interval (about 15 years) channel maintenance operations threaten riverine fishes in the Yazoo River basin despite growing recognition of the socio-economic and cultural value of this resource (Jackson et al., 1993).
We explicitly recognize that states are artificial geographic entities but
acknowledge that their jurisdictional and civic importance cannot be ignored
in any pragmatic approach to aquatic conservation in spite of our scientific
tendency (and necessity) to do so. Through statutory obligation, many state-bound,
natural-resource decision makers simply do not divide the landscape among natural
units, like drainages or watersheds, but see and manage only within political
boundaries. Even so, states are capable of recognizing problems and offering
solutions for recovery and management of fishes. For example, most southeastern
states now have heritage-based programs charged with inventorying and monitoring
(i.e., accounting information) imperiled fishes (e.g., Eager and Hatcher, 1980;
Mount, 1986; Warren et al., 1986; Terwilliger, 1991; Gilbert, 1992; Menhinick,
in press). State agencies also are making efforts to maintain fish diversity
rather than targeting specific species (e.g., Holman et al., 1993; Toth and
Aumen, 1994). Obversely, many state-based programs for nongame fishes are left
to languish on "soft" money (Williams, 1986; Pister, 1992), are underemphasized
(Cain, 1993; Angermeier and Karr, 1994), and lack the force of institutional
will or statutory authority, short of federal mandate, to effect change (Johnson,
1987; Pister, 1991).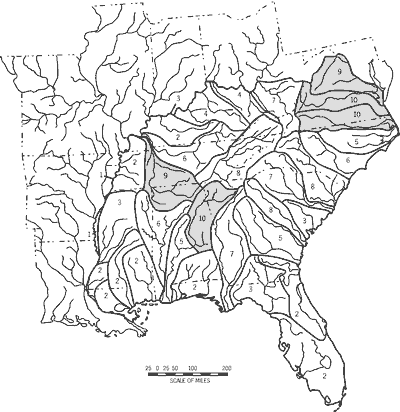
Notwithstanding parochial sovereignty and nongame fish program development in states, conflicts in use of streams among and within southeastern states are common (e.g., Voigtlander and Poppe, 1989; Neves and Angermeier, 1990; Anderson et al., 1991; Jackson et al., 1993; Saylor et al., 1993; Crawford, 1994). Of the 33 drainage units recognized here, 21 cross state boundaries (Figure 2). Obviously both federal-state and interstate cooperation toward long-term management of these drainages is appropriate and critical. Historical cooperation was limited to ostensible "improvement" of waterways for flood control and navigation, generally imposed through federal water projects, with incalculable losses to aquatic habitat (e.g., Hunt, 1988; Neves and Angermeier, 1990). Recent cooperation goes little beyond attempts to resolve acute and immediate water resource conflicts (e.g., Saylor et al., 1993) or occurs through federal abandonment of continued waterway "improvement" (e.g., Jackson and Jackson, 1989; Hupp, 1992). For example, the Tennessee River drainage includes portions of seven states, most of which are centers of fish imperilment and diversity. However, no comprehensive, coordinated management plan exists for the Tennessee, and the priority for sustainable management of this resource is low (Voigtlander and Poppe, 1989). Federal-state and intra- and interstate coordination is confounded within southeastern states primarily because jurisdiction over water, waterways, and the aquatic fauna is fragmented among agencies with different and often contradictory regulatory mandates (e.g., providing drinking water versus recreational fishing versus waste disposal).
If imperilment were the result of random sampling among families, familial imperilment would be proportional to familial representation in the total native fish fauna; however, this is not the case (Tables 3 and 4). Imperilment is disproportionately bestowed on both diverse and depauperate fish families. Fish families with disproportionately high levels of imperilment are characteristically dependent on vegetated, isolated wetland habitats; hypogean habitats; or benthic habitats (Tables 3 and 4). Three relatively depauperate fish families, the pygmy sunfishes (Elassomatidae), cavefishes (Amblyopsidae), and sturgeons (Acipenseridae), and two diverse families, darters and perches (Percidae) and bullhead catfishes (Ictaluridae) are exemplars. Imperiled members of the pygmy sunfishes primarily inhabit vegetated, often spring-fed, permanent wetlands (Rohde and Arndt, 1987; Mayden, 1993). These habitats, particularly those with springs, are among the most jeopardized in the Southeast (Etnier and Starnes, 1991; Cubbage and Flather, 1993; Mayden, 1993; Dickson and Warren, 1994). Likewise, the cavefishes depend on the integrity of a food base originating from surface waters and, ultimately, must live in a habitat that often serves as a sump for a variety of anthropogenic pollutants. Subterranean habitats and their dramatically evolved fishes are under threat worldwide (Groombridge, 1992). High imperilment also is concentrated among the sturgeons, darters and perches, and bullhead catfishes, all of which have a benthic life style. For some sturgeons, vulnerability to imperilment is associated with dams blocking migratory spawning runs, but most members of these families are dependent on the ecological integrity of the benthic environment and often require specific substrate sizes and configurations for spawning, feeding, and cover (Page and Swofford, 1984; Etnier and Starnes, 1993; Kessler and Thorp, 1993).
Table 5. Means and variation in range extent of native fishes grouped by status categories across 33 drainage units in the southeastern United States. Tabled entries include mean, two standard errors of the mean (2xSE), and coefficient of variation (CV). |
|||
Status |
Mean |
2 × SE |
CV |
AFS-E (n = 14) |
3.31 |
1.34 |
80 |
AFS-T (n = 36) |
4.91 |
1.86 |
114 |
AFS-SC (n = 34) |
5.91 |
2.04 |
103 |
AFS-ETSC (n = 84) |
5.01 |
1.16 |
108 |
No Status (n = 409) |
7.7 |
0.76 |
99 |
1 Values not significantly different in orthogonal contrasts. |
|||
High levels of imperilment in fishes with multiple niche axes converging on benthic resources is not unexpected. Degradation of streams is often first manifested in benthic habitats and communities (Reice and Wohlenberg, 1993). Streams entrain organic and inorganic material from the watershed, and this material is deposited, stored, and biologically recycled largely on or within the streams substrate (Merritt et al., 1984). Being in intimate contact with these materials, benthic fishes (and their benthic food resources) are affected directly by sediment particles (Minshall, 1984; Berkman and Rabeni, 1987; Haro and Brusven, 1994) and by nutrients and toxins contained within sediments (Reice and Wohlenberg, 1993). We conclude that the disproportionately high imperilment of these benthic fish families is simply the initial, expected manifestation of long-term, complex, cumulative aquatic degradation.
Mean range extent was lower for imperiled than nonimperiled fishes (Table 5), and probability of imperilment increased with decreasing range extent. However, the explanatory power of these relationships is weak. Although significant statistically, conservation status accounted for only 2.2 percent of the variation in range extent in the analysis of variance model. Likewise, in the logistic regression model, the increase in probability of imperilment as range extent decreases was low, and misclassifications of imperiled and nonimperiled fishes was high.
Table 6. Means and measures of spatial variation for fish taxa richness, unique taxa, imperilment, and stream-type diversity for 33 drainage units of the southeastern United States. Table entries each include mean, two standard errors of the mean (2 ? SE), minimum (Min.) and maximum (Max.) values, coefficient of variation (CV), Index of Dispersion (ID), and the significance level (p-value) for departure from a random spatial distribution. |
|||||||
Variable |
Mean |
2 × SE |
Min. |
Max. |
CV |
ID |
p-value |
Fish Taxa Richness |
108.6 |
10.97 |
56 |
193 |
29 |
292 |
p<< 0.01 |
Unique Taxa |
3.9 |
1.44 |
0 |
16 |
107 |
142 |
p<< 0.01 |
Imperilment |
|
|
|
|
|
|
|
AFS-ET |
3.2 |
0.59 |
0 |
14 |
104 |
113 |
p<<0.01 |
AFS-ETSC |
6.2 |
0.39 |
1 |
19 |
83 |
138 |
p<<0.01 |
Stream Types |
5 |
1 |
1 |
10 |
57 |
53 |
p<0.05 |
Imperilment, a measure of extinction potential, and range extent, a categorical measure of area inhabited, might be expected to show stronger relationships than detected in these models. Restricted geographic range is often associated with fish vulnerability to extinction (Moyle and Williams, 1990; Etnier and Starnes, 1991; Angermeier, 1995). The weak explanatory power of the resultant models is less enigmatic considering:
Many nonimperiled fishes in the Southeast are restricted to one or few drainage units (i.e., limited in range extent) but occur widely and commonly within those units. In contrast, some imperiled fishes are known from a number of southeastern drainage units but inhabit very limited stream reaches within each unit. Populations of the latter are effectively subject to effects of reduced habitat area and insularization (Angermeier, 1995) regardless of overall range extent. In either case, our measure of range extent does not account well for the actual area occupied. If examined at finer spatial scales (e.g., kilometers of stream inhabited), geographic range might account for greater variance in imperilment, and limited range might show a higher increase in risk of imperilment. Even at smaller spatial scales and using only extirpated fishes, Angermeier (1995) also noted relatively weak statistical associations between extinction and limited physiographic range for fishes in Virginia.
Table 7. Results of correlation analyses (Pearson's correlation coefficient and Kendall's tau-b coefficient) of fish imperilment and drainage-unit area, native fish taxa richness, unique taxa, and stream-type diversity in 33 drainage units of the southeastern United States. AFS-ET and AFS-ETSC refer to combinations of American Fisheries Society conservation status categories (see Williams et al., 1989). |
||||
Status Category |
Drainage-unit Area1 |
Fish Taxa Richness |
Unique Taxa |
Stream-type Diversity |
AFS-ET |
||||
|
0.374
|
0.753
|
0.800
|
0.401
|
|
0.214
|
0.589
|
0.363
|
0.177
|
AFS-ETSC |
||||
|
0.448
|
0.828
|
0.854
|
0.425
|
|
0.249
|
0.636
|
0.47
|
0.247
|
1 n = 30. |
||||
Weak relationships between range extent and imperilment are expected if widespread, complex degradation of southeastern fish habitats is a strong determinant of imperilment (Angermeier, 1995). Complex degradation over a large geographic area involves numerous forms of insult to aquatic systems that affect various vulnerable fish taxa differentially depending on the intensities and combinations of exposures. Angermeier (1995) identified three uncontrolled factors interacting to randomize degradation "treatments" among species in ecosystems subject to anthropogenic impacts:
Table 8. Summary of multiple regression analyses of fish imperilment, drainage-unit area, native fish taxa richness, stream-type diversity (S-T) , and unique taxa (UT) in 33 drainage units of the southeastern United States. Tabled entries include intercept, slope, and the proportion of variance (R2) in imperilment accounted for by each model. AFS-ET and AFS-ETSC refer to combinations of American Fisheries Society conservation status categories (see Williams et al., 1989). * = probability (parameter = 0) < 0.10; ** = probability (parameter = 0) < 0.05; *** = probability (parameter = 0) < 0.001. |
||||||||||
|
|
Model 1 |
Model 2 |
||||||||
Status Category |
Intercept |
Richness |
S-T |
UT |
Area |
R2 |
Intercept |
Richness |
S-T |
R2 |
AFS-ET |
-4.46** |
1.99*** |
0.04* |
-0.11 |
0.18 |
0.616 |
-3.43*** |
1.85*** |
0.03** |
0.609 |
AFS-ETSC |
-5.65*** |
2.51*** |
0.04** |
-0.11 |
0.23 |
0.809 |
4.39*** |
2.38*** |
0.04*** |
0.800 |
Fish taxa richness, unique taxa, and stream-type diversity showed variable, but aggregated, distribution patterns that overlap considerably with one another and with the spatial distribution of imperilment in drainage units of the Southeast (Table 6; Figures 4-8). Statistical results generally supported association of these attributes and fish imperilment (Table 7). The Tennessee-Cumberland Region (DU26-DU28) and Mobile Bay Basin Region (DU16-DU18) consistently had one or more drainage units forming part of a core of high fish taxa richness, unique taxa, and imperilment (Figures 4-6). Drainage units adjacent to these core regions usually showed intermediate values (i.e., Southeastern Ohio River Region, DU29-DU33). Most drainage units of the Atlantic and Gulf slopes and Mississippi embayment showed low to intermediate values for fish taxa richness, unique taxa, and imperilment.
Drainage units congruent in levels of taxa richness, unique taxa, or stream-type diversity and imperilment often share common biogeographic histories. Although the relationships between imperilment and these other attributes were positive (Table 7), we discern a clear temporal distinction in the origin of imperilment and the other attributes. Patterns of fish taxa richness, unique taxa, and stream types in the Southeast all have a strong association with Earth history (Burr and Page, 1986; Hocutt et al., 1986; Starnes and Etnier, 1986; Swift et al., 1986; Mayden, 1988; Warren et al., 1991). In contrast, patterns of imperilment are primarily products of the past century that were only documented in the past 30 or so years (Deacon et al., 1979; Miller et al., 1989; Williams et al., 1989). For example, the highlands of the Tennessee-Cumberland, Southeastern Ohio River, and the Mobile Bay Basin regions were subjected to a series of complex, major geological vicariance events initiated in the pre-Pleistocene and as a result, were relatively isolated from much of the remaining diverse and widespread Central Highlands ichthyofauna (Mayden, 1988). Likewise, much of the ichthyofaunal composition and endemicity of the Atlantic and Gulf slope lowlands is associated with eustatic cycles, dating at least to the Miocene, superimposed on physiographic features and drainage trends (Swift et al., 1986). The implication of the association of imperilment with recent environmental degradation following millions of years of Earth history is clear. In a geological "blink of the eye" the fish fauna across the entire Southeast is being compromised.
The largest increases in imperilment of fishes overlapped with high fish taxa richness, unique taxa, and stream-type diversity, but substantial increases also occurred across a wide diversity of drainage units (e.g., Everglades-Tampa Bay-Waccasassa River Drainages, DU12, and Mississippi River Mainstem, DU25) (Figure 7). However, even at relatively low rates of increase, imperilment of the fauna can increase dramatically in brief time periods. A 0.2 percent annual increase in imperilment in a given drainage unit for 50 years yields a fauna with nearly ten percent of its members imperiled. We emphasize our estimates of increases in imperilment for southeastern drainages are uncertain and not amenable to tests for accuracy or precision, particularly for predictive purposes. We do not suggest by the hypothetical example that even low rates of imperilment or more correctly, its corollary, extinction, could be sustained over long periods (e.g., 100 years? 200 years?). The numbers of imperiled and ultimately, extinct fishes, necessarily reach an asymptote as the pool of vulnerable taxa is exhausted over time. Nevertheless, it is certain that integrity of the fish fauna will not be maintained even if low rates of increase are projected into the next century.
The mapped patterns of imperilment, increased imperilment, fish taxa richness, and unique taxa among southeastern drainage units provide a starting point for applying conservation triage and prioritizing proactive efforts to sustain overall fish and other aquatic diversity within a historical ecological context (Figures 3-7). Certain geographic aggregations of drainage units are in greater need of action than others in terms of sheer numbers of imperiled, unique, or native taxa, particularly the Tennessee-Cumberland and Mobile Bay Basin regions. We note, however, that every drainage unit in the Southeast shows some level of imperilment and many show high levels of increase in imperilment regardless of fish taxa richness or stream-type diversity. The management actions required to go beyond triage or simple prioritization and actually affect change must be grounded in a framework of interstate and federal cooperation that is, to date, unprecedented.
Fish Taxa Richness and Stream-Type Diversity
Drainage units with high fish taxa richness and stream-type diversity contained more imperiled species than units with low richness and stream-type diversity (Table 8). In the final regression models, however, fish taxa richness accounted for 85 percent of the Type II SS contributed by fish taxa richness and stream-type diversity. The predictive power of the regression models thus lies primarily in fish taxa richness; national patterns of animal endangerment are associated similarly (Flather et al., 1994). A log-log regression model of imperilment and fish taxa richness approximates a power function (Conner and McCoy, 1979), where: I = k(R)Z and I is imperilment, R is fish taxa richness, and k and Z are constants. Interpreted as such, the proportion of imperiled fishes in a drainage unit increases with increasing fish taxa richness. This does not imply direct causation or circularity (e.g., high taxa richness begats high imperilment). Rather, a hypothesis is supported that imperilment (as a measure of extinction vulnerability) is largely an epiphenomenon of sampling of the available pool of fish taxa in a drainage unit. Similar hypotheses are prescribed for analogous regression models (Connor and McCoy, 1979; Angermeier and Schlosser, 1989). Simply, drainage units with large fish faunas have more individual fish taxa vulnerable to extinction. This result is a direct expectation under a scenario of complex, pervasive degradation of aquatic habitats (Angermeier, 1995). We conclude that conditions favorable for extinction are pervasive across southeastern drainages but are most visibly manifested in the richest drainage units.
Drainage-Unit Area
Imperilment is not predicted effectively by drainage-unit area despite correlation of drainage area and fish taxa richness in subsections of the Southeast (Swift et al., 1986; Sheldon, 1987; Warren and Angermeier, in press). Drainage unit area not only failed to add explanatory power to multiple regression models (Table 8), but it was not an effective substitute for total native taxa richness in predicting imperilment. As a single independent variable, drainage-unit area explained less than 15 percent of the variation in imperilment. From this, we infer two things:
The relative lack of usefulness of unique taxa for predicting imperilment complements the weak associations between range extent and imperilment. In addition, we found no difference for the total native fish fauna in the proportion of unique taxa among imperiled and nonimperiled fishes (LI = 2.58, p < 0.1337). We do not suggest these results negate the intuitively appealing and empirically supported associations between restricted geographic range and imperilment in fishes (Etnier and Starnes, 1991; Moyle and Leidy, 1992; Angermeier, 1995) or other organisms (Flather et al., 1994). Instead, we invoke reasoning analogous to that forwarded for the models of range extent and imperilment. Vulnerability to extinction in southeastern fishes is not a simple function of endemicity (or "degree" of cosmopolitanism). Some unique taxa are widely distributed within a drainage unit; others are severely limited in distribution. Via the randomized "treatments" of pervasive, complex degradation, vulnerability to extinction may be allocated among fishes with diverse range sizes and ecological attributes (Angermeier, 1995). Weak associations of range size and imperilment in concert with the final multiple regression models support a scenario of pervasive, complex habitat degradation as a strong determinant of observed imperilment patterns in southeastern fishes.
Development of models associating vulnerability to the extinction process and ecological and zoogeographic characteristics of organisms and communities is a high priority for conservation biology (Soulé and Kohm, 1989). Aside from this effort, few related analyses are available for fishes in the Southeast (Sheldon, 1987; Nagel, 1991; Angermeier, 1995). Geographic and ecological range restrictions are primary among attributes associated with many southeastern imperiled, extirpated, and extinct fishes (Mayden, 1992; Etnier and Starnes, 1991; Angermeier, 1995). We find that several promising geographic and faunal attributes (i.e., drainage-unit area, range extent, and unique taxa) are not completely satisfactory in explaining individual fish imperilment nor levels of imperilment in drainage units, but this finding is not unprecedented (Moyle and Leidy, 1992; Frissel, 1993; Etnier and Starnes, 1991; Angermeier, 1995). We do not construe this as meaning no associations exist among imperilment and these geographic and faunal attributes but only that no strong associations exist (Angermeier, 1995).
The final models identifying taxa richness, and to a lesser degree, stream-type diversity as the best predictors of imperilment were more revealing and complementary to the relative ineffectiveness of other variables. The synthesis of these results implicates pervasive, complex degradation of fish habitats across southeastern drainages as the engine of imperilment. To this we add an important caveat taken from the familial analyses. Degradation appears most strongly manifested in imperilment of fishes associated with benthic habitats; a habitat predictably impacted first by cumulative, long-term abuse of aquatic systems. Decline of other benthic organisms, such as freshwater mussels, strongly supports this contention (Williams et al., 1993).
Society must recognize that all upstream activities, including those on the terrestrial component of the catchment, have cumulative downstream effects and address this reality in prioritization, resource allocation, and implementation of conservation management. The linear, unidirectional nature of rivers and streams is a singular attribute that dictates emphasis on whole-system approaches to management. The prevailing strategy of waiting for species to become imperiled before activating conservation programs is a major and costly shortcoming in conservation in the Southeast and elsewhere (Scott et al., 1988; Blockstein, 1992; Angermeier, 1995). The current situation for aquatic resource management among and within southeastern states is described succinctly by Noss and Cooperrider (1994; page 264): "Nowhere is the fragmentation of our thinking and institutions and the arrogance of our management more pronounced than in our stewardship of aquatic ecosystems."
We believe it should be quite clear that successful management for aquatic biological integrity must transcend political boundaries, jurisdictional subdivisions, and short-term economic policies and ultimately focus on the long-term interaction of humans and the environment within ecologically definable units (Karr, 1993; Angermeier and Karr, 1994; Maser and Sedell, 1994; Noss and Cooperrider, 1994). Society must move toward proactive management centered on maintaining ecological integrity of aquatic ecosystems and hence protecting existing diversity as a whole (Blockstein, 1992; Hughes and Noss, 1992; Allan and Flecker, 1993; Angermeier and Karr, 1994; Marcot et al., 1994).
The last line of defense against extinction of fishes in the Southeast and elsewhere in the United States is the Endangered Species Act of 1973, as amended. It should remain just that, the last line of defense. Clearly, this Act, the strongest environmental law on Earth, cannot begin to meet the herculean task of conserving the Southeasts imperiled fishes, and as a species-by-species safety net, it simply cannot and should not be expected to function alone in conservation of the great southeastern fish fauna. We need shifts in management approaches that avert continued endangerment of fishes. The foundation of such an approach should include a system-led (e.g., drainage unit) rather than species-led focus; explicit biological integrity goals in the context of preventing degradation of high-quality systems and restoring poor-quality systems; commitment to implementing effective land-water management practices rather than implementing bureaucracies; and recognition of land and water resources as integrated parts of the same system.
We present evidence from the fish fauna pointing to the widespread, pervasive decline of aquatic habitats across the Southeast. The associated problems, if there is a will to correct them (see Meyer, 1995), are simply beyond the statutory and fiscal abilities of any one piece of legislation or agency to correct. In short, we believe the need for transcendence, focus, and action is clear and urgent.